
Quantitation of the immunodominant 33-mer peptide from α-gliadin in wheat flours
In wheat there are multiple fractions able to activate the adverse response of the human immune system. Among these fractions the most active is that called 33-mer because it is the most resistant to human digestion and because it contains six copies of the three toxic epitopes and its intermolecular bonds are very strong. It is therefore important to know the quantity of this fraction in the grains. The study of which some parts are reported, examined 57 different types of wheat, ancient and modern, noting that the difference, in all soft wheat and spelt flour, of 33-mer is wide: from 90.9 to 602.6 μg / g made with flour. On the other hand, its presence in monococcum wheat and durum wheat was not detected. These results take on great importance because they allow grains to be chosen with limited or no presence of this important toxic fraction for products that are more suitable for non-celiac gluten sensitive people or those suffering from gluten disorders.
“All gluten protein fractions, namely the alcohol-soluble prolamins and the insoluble glutelins, contain CD-active epitopes3. The prolamin fraction is particularly rich in proline and glutamine and the numerous proline residues lead to a high resistance to complete proteolytic digestion by human gastric, pancreatic, and brushborder enzymes. Studies by Shan et al. (2002) showed that a large 33-mer peptide (LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF) from α2-gliadin (position in the amino acid sequence of α2-gliadin: 56–88) is resistant to cleavage by intestinal peptidases4,5. The 33-mer is widely called the most immunodominant gluten peptide4,6,7, because it contains three overlapping T-cell epitopes, namely PFPQPQLPY (DQ2.5-glia-α1a, one copy), PYPQPQLPY (DQ2.5-glia-α1b, two copies) and PQPQLPYPQ (DQ2.5-glia-α2, three copies)3, which result in the initiation of a strong immune response.
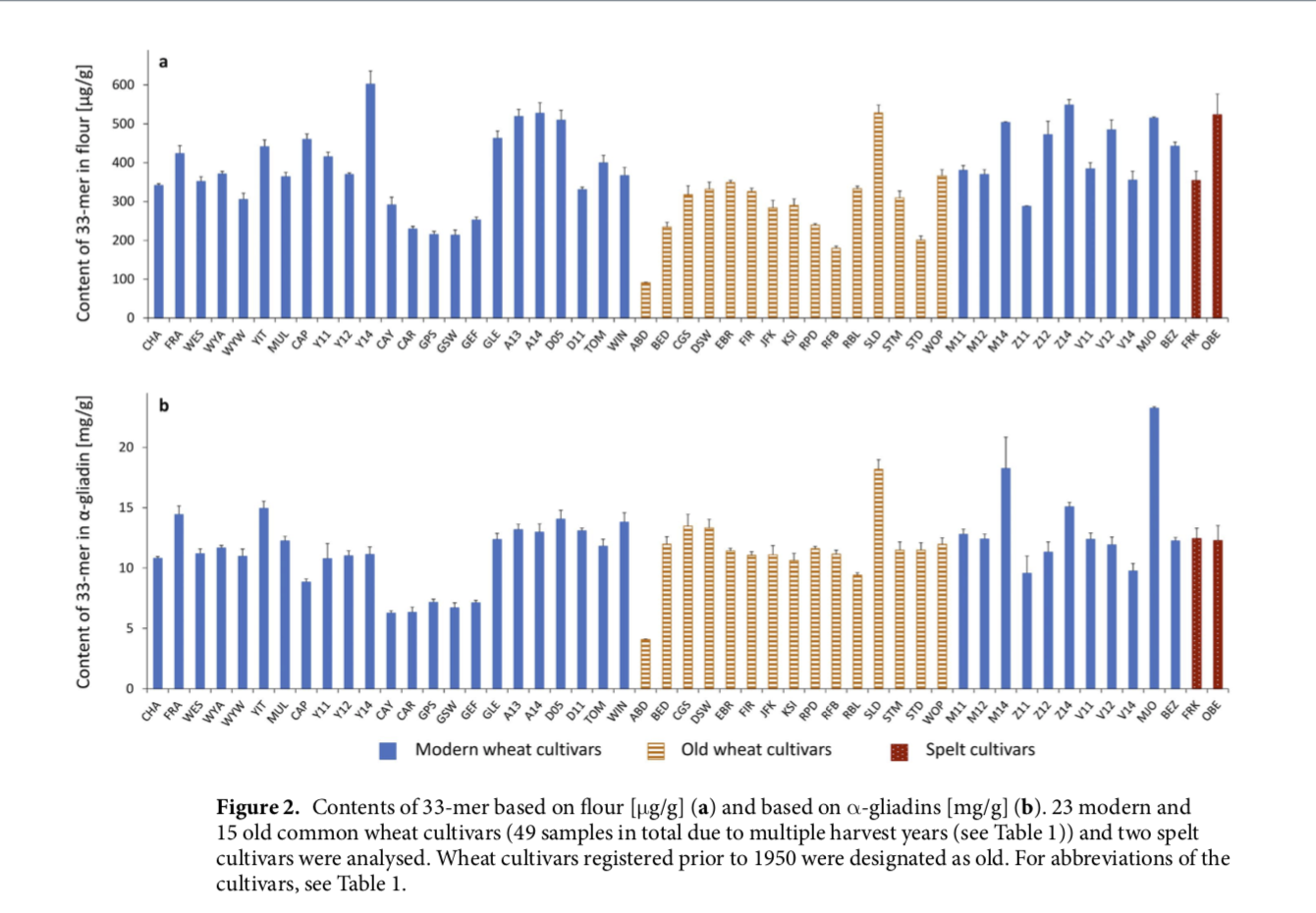
The high relevance of the 33-mer is reflected by the production of two monoclonal antibodies (A1 und G12) against the 33-mer peptide18. These are used in commercially available enzyme-linked immunosorbent assays (e.g., GlutenTox ELISA, Biomedal, Sevilla, Spain and AgraQuant® ELISA Gluten G12, Romer Labs, Tulln, Austria) for the immunochemical quantitation of gluten in supposedly gluten-free foods19. Although about 20 papers per year were published since 2000 and 53 for the year 2016, information about the quantities of 33-mer in different wheat species and cultivars is still missing. The amount of 33-mer was determined in 57 samples of different wheat species from around the world (Table 1), including hexaploid common wheat (T. aestivum) and spelt (T. aestivum ssp. spelta), tetraploid durum wheat (T. turgidum durum) and emmer (T. turgidum dicoccum), and diploid einkorn (T. monococcum) to make a precise assessment of the importance of this peptide associated with CD. The 33-mer was present in all common wheat and spelt flours in a range from 90.9 to 602.6 μg/g of flour (Fig. 2a). The modern wheat Y14 had the highest amount of 33-mer (602.6 μg/g flour), that was significantly dif- ferent to all analysed cultivars with the exception of Z14 (see Supplementary Table S2). The old wheat ABD with the lowest 33-mer content of 90.9 μg/g flour differed significantly to all wheat and spelt cultivars. In contrast, the old wheat SLD (528.0 μg/g flour) contained one of the highest 33-mer amounts of the analysed flours and showed no significant difference to the modern wheats A13, A14, D05, Z14, V12, and MJO and spelt OBE. Special atten- tion was directed to MJO, because the 33-mer was first identified in this cultivar13. The content of 33-mer in MJO (515.0 μg/g flour) showed no significant difference to A13, A14, CAP, D05, GLE, Z12, Z14, V12, and OBE. Most of the modern and old wheat flours contained the 33-mer in a range of 200–400 μg/g flour with an overall average of 368 ± 109 μg/g flour. As a result, only some differences in 33-mer contents between these wheat cultivars were significant. A certain trend, e.g., that modern wheat cultivars generally contain higher amounts of 33-mer than old cultivars could not be derived from the data. Considering the amounts of 33-mer in the two spelt cultivars, it was noticeable that OBE contained one of the highest amounts of the 33-mer peptide (523.4 μg/g flour). The content of 33-mer in FRK (353.9 μg/g flour) was in the range of 200–400 μg/g flour, and did not differ significantly from the common wheat cultivars. Influence of harvest year and cultivar on the contents of 33-mer. To see whether harvest year or cultivar had a greater influence on 33-mer contents, four wheat cultivars (Mv Magvas, Mv Mazurka, Mv Verbunkos, and Yumai-34) grown at the same location in Hungary (Martonvásár) and harvested in three years (2011, 2012, and 2014)35 were studied. The harvest year significantly influenced the 33-mer contents (p < 0.001), whereas the cultivars did not (p = 0.391). There were no significant differences in 33-mer contents between the four cultivars within the harvest year 2011, two out of six differences (V12 vs. M12 and V12 vs. Y12) were signif- icant (p < 0.05) within the harvest year 2012, and three out of six (V14 vs. Z14, V14 vs. Y14, and V14 vs. M14) within the harvest year 2014. Apparently, the environmental factor had a greater influence on 33-mer contents than the genetic background of the four wheat cultivars, because the results for each combination of harvest years (2011 vs. 2012, 2011 vs. 2014 and 2012 vs. 2014) were significantly different (p ≤ 0.034). Analysis of durum wheat, emmer and einkorn. The 33-mer peptide was also analysed in two durum wheat and two emmer cultivars (genome AABB) as well as two diploid einkorn cultivars (genome AA) (Table 1). In each of these wheat species, the 33-mer was not detected (<LOD). In comparison to hexaploid common wheat, durum wheat, emmer, and einkorn do not contain the D-genome, which originated from hybridisation of T. turgidum dicoccum (genome AABB) with Aegilops tauschii (genome DD)36. The absence of the 33-mer pep- tide can be explained by the fact that this peptide is encoded by genes located in the Gli-2 locus on chromosome 6D, which is missing in durum wheat, emmer, and einkorn. Studies by Molberg et al. showed clear variations in intestinal T-cell responses between common wheat and tetra- or diploid species due to different degrees of T-cell immunoreactivity between the gluten proteins encoded on the A-, B-, and D-genome. Einkorn cultivars were only recognized by DQ2.5-glia-α1a-specific T-cell clones, but not by DQ2.5-glia-α1b- and DQ2.5-glia-α2-specific T-cell clones. Emmer and durum wheat cultivars were all recognized by DQ2.5-glia-α1a-specific T-cell clones, but only two out of four emmer cultivars and three out of ten durum wheat cultivars activated DQ2.5-glia-α1b- and DQ2.5-glia-α2-specific T-cell clones37. Consistent with our results, Prandi et al.38 found that the 33-mer was not present in durum wheat. As a consequence, this peptide was used as a marker peptide to identify the presence of common wheat in durum wheat flours. One durum wheat cultivar was also analysed by van den Broeck et al.33 and the 33-mer peptide was not detected either. Discussion The present study is the first to establish a SIDA combined with targeted LC-MS/MS for the quantitative deter- mination of the immunodominant 33-mer peptide in wheat flours. Due to the use of a stable-isotope-labelled *33-mer standard, sample preparation could be simplified without reduction/alkylation and SPE purification. Although the UniProtKB database had only 19 out of 897 entries for α-gliadin sequences from Triticinae con- taining the 33-mer with an identity of 100%, all 40 analysed modern and old common wheat and spelt cultivars contained the immunodominat 33-mer peptide (51 flour samples in total, because several flours were available from different harvest years). The focus on this peptide seems to be legitimated not only because of its unique structure containing six copies of three overlapping T-cell epitopes, but also because of its presence in all hexa- ploid wheat cultivars analysed in this study. PCA analysis of the data demonstrated that the contents of 33-mer were not suitable to differentiate old from modern common wheat cultivars, because cultivars with high 33-mer contents were found within both flour sets. The 33-mer was not detected in two cultivars each of tetraploid emmer and durum wheat as well as diploid einkorn, which do not contain the D-genome. This observation may be explained by the fact that the 33-mer is encoded on the Gli-2 locus on chromosome 6D, but a larger set of durum wheat, emmer and einkorn cultivars would have to be analysed to conclude whether these wheat species generally lack the 33-mer peptide. Further work will focus on correlating the content of 33-mer analysed by LC-MS/MS with the gluten content determined by ELISA using the G12 monoclonal antibody. Quantitation of the immunodominant 33-mer peptide from α-gliadin in wheat flours 2017. Kathrin Schalk, Christina Lang, Herbert Wieser, Peter Koehler & Katharina Anne Scherf . Scientific Reports.”
The study “Differentiation between Celiac Disease, Nonceliac Gluten Sensitivity, and Their Overlapping with Crohn’s Disease: A Case Series. Aristo Vojdani and David Perlmutter. December 2012 “ showed the reaction of the immune system of a non-celiac gluten sensitive patient to some fractions of the gliadin including the 33-mer fraction. Although there is no enough data concerning the link between the subunits of gliadin and the NCGS, there is no doubt that the peptides that are more resistant to human digestion are the ones most involved: and the 33-mer is, in this “classification” the number 1.
Back