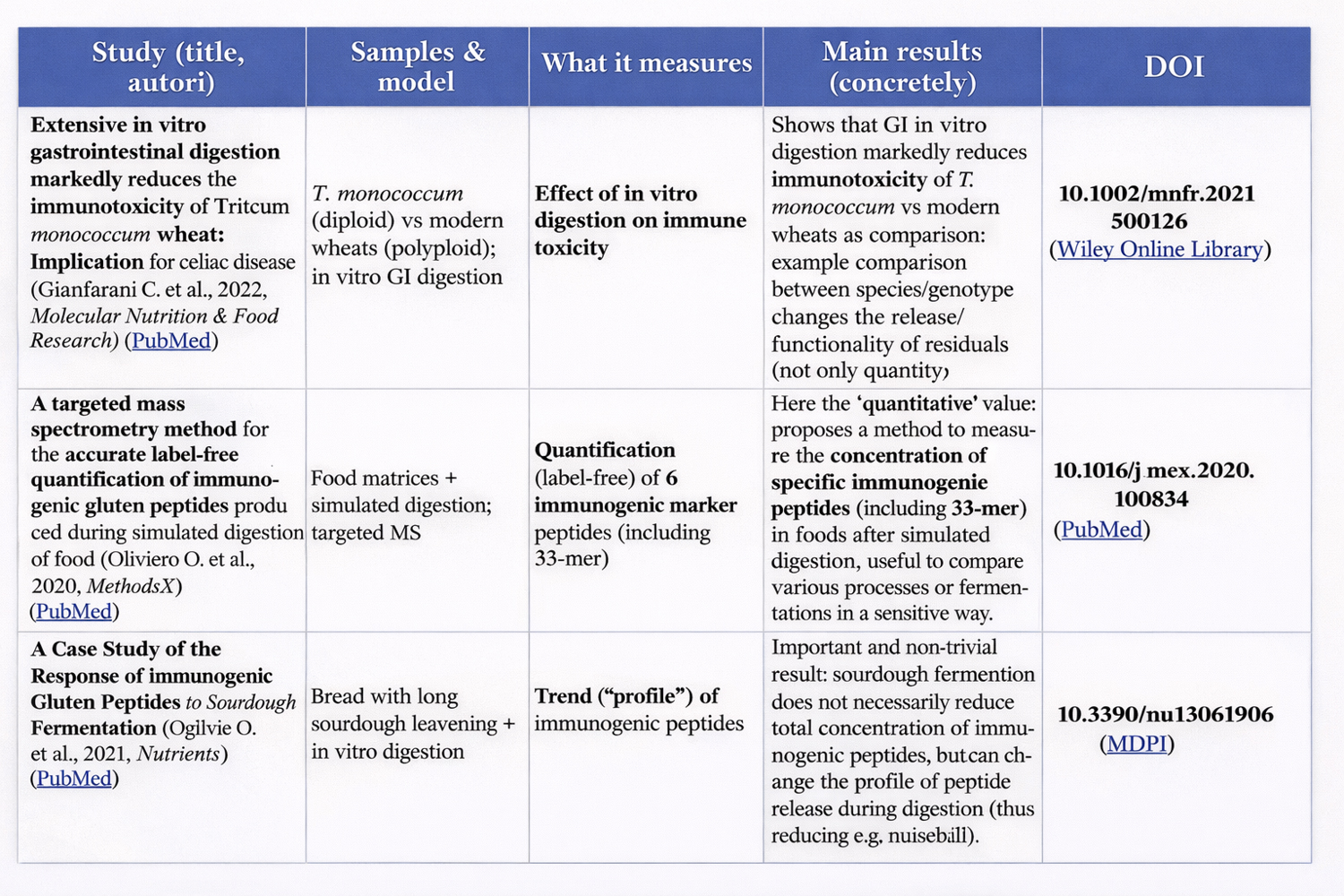
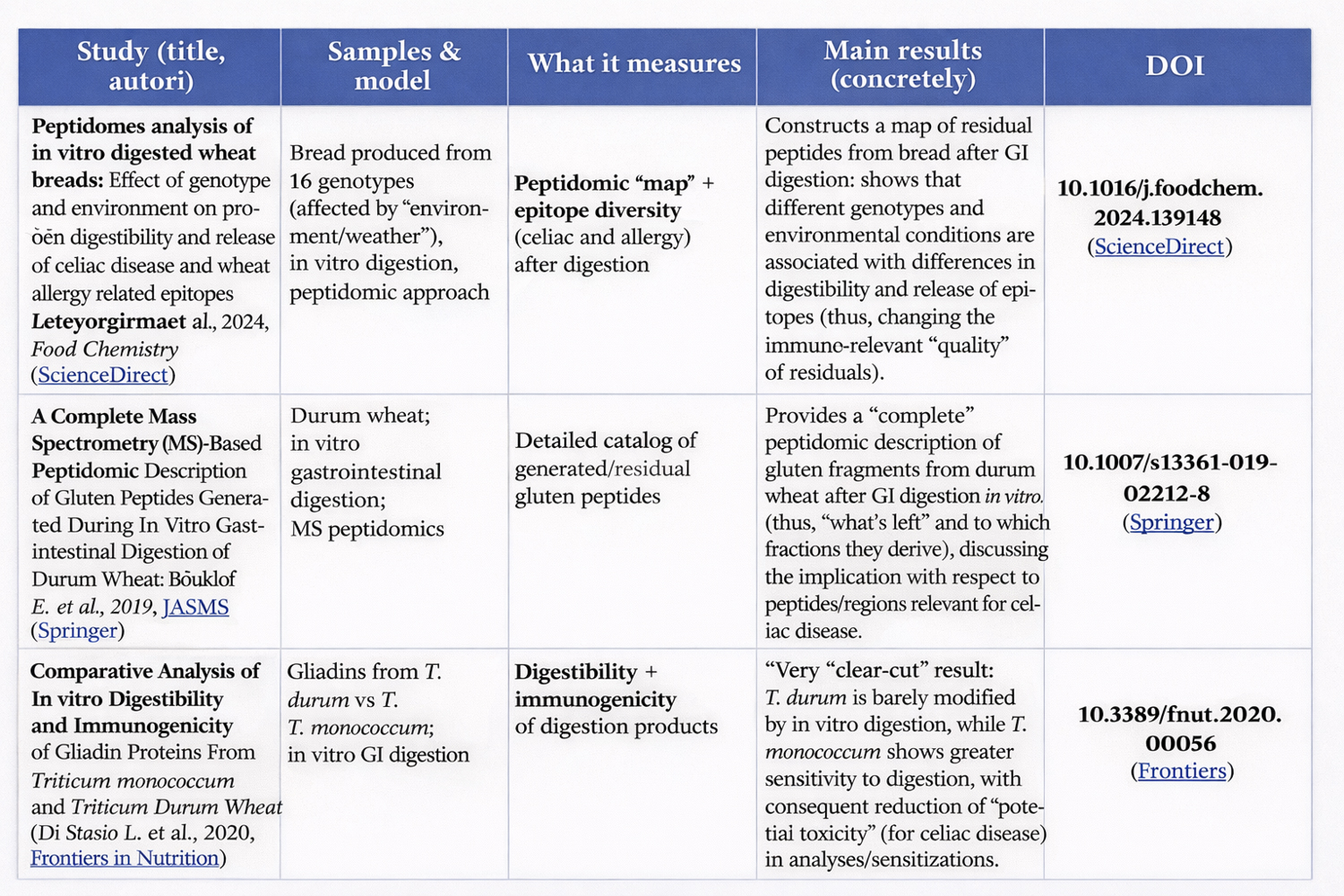
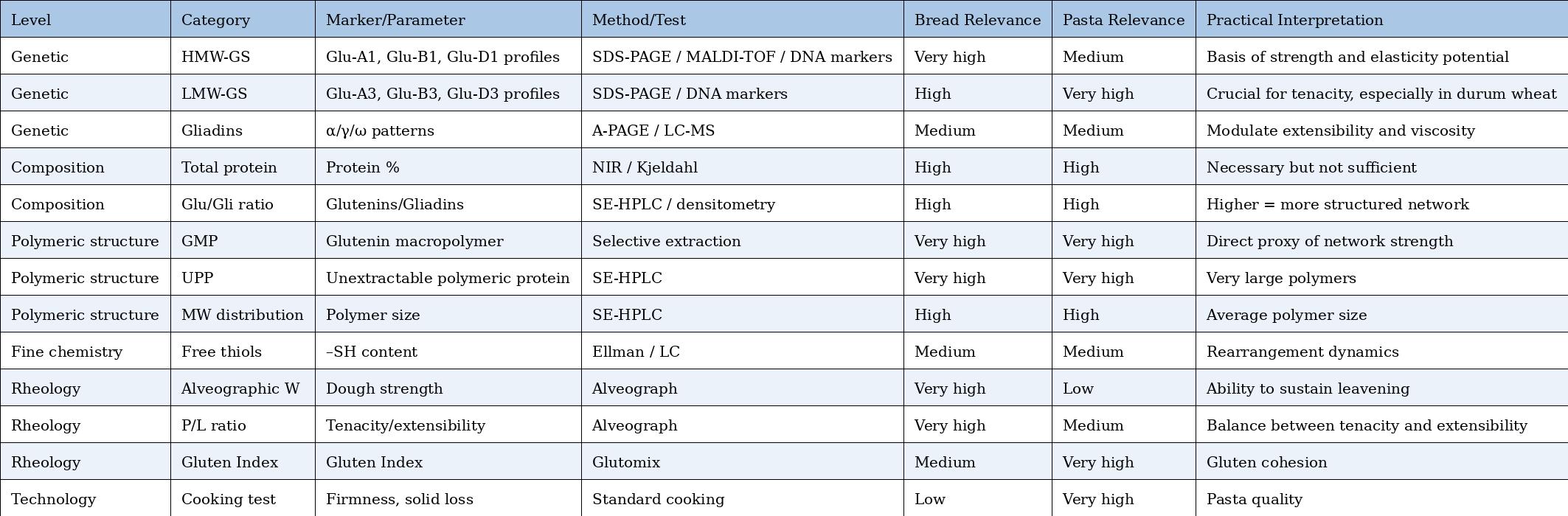
✅ RELATED ARTICLE 1
Mitochondria and Oxidative Stress
Highlight
Mitochondria are the main source of ROS in the body and at the same time one of the primary targets of oxidative damage.
Their efficiency largely determines the level of cellular oxidative stress.
What mitochondria do
Produce ATP via oxidative phosphorylation
Regulate apoptosis
Participate in cellular signaling
Regulate nutrient metabolism
During energy production, a small fraction of electrons escapes from the respiratory chain, forming superoxide.
Why mitochondria produce ROS
In the electron transport chain:
O₂ + electron → O₂•⁻
This is a physiological and unavoidable event.
BOX — Physiological production
A moderate production of mitochondrial ROS is necessary for:
adaptive signaling
Nrf2 activation
mitochondrial biogenesis
What is mitochondrial dysfunction
A condition in which:
ATP production decreases
electron leakage increases
ROS production increases
A vicious cycle is created:
Inefficient mitochondrion → more ROS → mitochondrial damage → even less efficient mitochondrion
Factors that damage mitochondria
chronic hyperglycemia
excess oxidized fats
inflammation
toxins
micronutrient deficiencies
sleep deprivation
Mitochondria and chronic diseases
Mitochondrial dysfunction observed in:
type 2 diabetes
cardiovascular disease
neurodegeneration
sarcopenia
aging
How to improve mitochondrial function
Nutrition
adequate protein intake
micronutrients (B vitamins, iron, copper, magnesium)
polyphenols
Physical activity
aerobic exercise
resistance training
Sleep
regularity
7–9 hours
Stress
reduction of chronic load
BOX — Key concept
Oxidative stress is not reduced by “turning off ROS.”
It is reduced by making mitochondria more efficient.
Conclusion
The mitochondrion is the central hub of redox metabolism.
Protecting mitochondrial function means acting upstream on oxidative stress.
✅ RELATED ARTICLE 2
Circadian Rhythm and Oxidative Stress
Highlight
The circadian rhythm coordinates the expression of genes involved in metabolism, energy production, and antioxidant systems.
When this timing system is altered, ROS production increases and the capacity to neutralize them decreases, promoting chronic oxidative stress.
What is the circadian rhythm
A biological timing system of about 24 hours that regulates:
sleep–wake cycle
hormone secretion
energy metabolism
body temperature
cellular repair activity
The main control center is the suprachiasmatic nucleus of the hypothalamus, mainly synchronized by light.
Central and peripheral clocks
There are:
one central clock (brain)
peripheral clocks (liver, muscle, pancreas, adipose tissue, heart)
These clocks regulate the temporal expression of thousands of metabolic genes.
BOX — Key concept
Not only what you do, but also when you do it influences redox metabolism.
Link between circadian rhythm and antioxidant systems
Many antioxidant enzymes show circadian oscillations:
superoxide dismutase (SOD)
catalase
glutathione peroxidase
Glutathione synthesis also follows a daily rhythm.
If rhythm is disturbed, these oscillations flatten → lower antioxidant defenses.
Circadian rhythm and mitochondria
The biological clock regulates:
mitochondrial biogenesis
fusion/fission dynamics
respiratory chain efficiency
Circadian misalignment → less efficient mitochondria → greater electron leakage → more ROS.
What disrupts circadian rhythm
evening artificial light
nighttime screen exposure
shift work
insufficient sleep
irregular or nighttime meals
social jet lag
Biological effects of misalignment
Chronic misalignment causes:
increased ROS production
reduced antioxidant activity
increased inflammation
altered glucose and lipid metabolism
BOX — Simplified mechanism
Altered rhythm → inefficient mitochondria → ↑ ROS
Altered rhythm → ↓ antioxidant enzymes
Result: oxidative stress
Circadian rhythm and chronic diseases
Associated with higher risk of:
obesity
type 2 diabetes
metabolic syndrome
cardiovascular disease
cognitive decline
Partly through increased systemic oxidative stress.
Sleep: the main redox “reset”
During sleep:
brain metabolism decreases
antioxidant activity increases
DNA repair systems activate
mitochondrial efficiency improves
Sleep deprivation → measurable increase in oxidative stress markers after only a few nights.
Meal timing and oxidative stress
Eating at biologically inappropriate times:
worsens glycemic control
increases mitochondrial ROS production
promotes lipotoxicity
An eating window aligned with the light–dark cycle improves redox balance.
How to protect circadian rhythm
Light
natural light in the morning
reduced blue light in the evening
Sleep
regular schedule
adequate duration
Meals
consistent timing
avoid large nighttime meals
Physical activity
preferably during daytime
BOX — Key concept
Without a functional circadian rhythm, even a perfect diet and good supplements have limited effectiveness on oxidative stress.
Integration with other pillars
Circadian rhythm acts in synergy with:
mitochondrial function
exercise
stress management
Protecting rhythm is a primary lever in oxidative stress prevention.
Conclusion
The circadian rhythm is a fundamental regulator of redox balance.
Its disruption promotes both increased ROS production and reduced antioxidant defenses, creating conditions for chronic oxidative stress.
Preserving the light–dark rhythm is one of the most powerful and underestimated interventions for cellular health.
✅ RELATED ARTICLE 3
Exercise, Hormesis, and Nrf2: Why Movement Reduces Oxidative Stress
Highlight
Exercise transiently increases ROS production, but this controlled stimulus activates powerful adaptive mechanisms that enhance endogenous antioxidant defenses.
This phenomenon is known as hormesis and is largely mediated by the transcription factor Nrf2.
The exercise paradox
During physical activity:
oxygen consumption increases
mitochondrial electron flux increases
ROS production temporarily increases
Yet, in the long term, regularly trained individuals show lower basal oxidative stress.
BOX — Apparent paradox
Exercise produces ROS, but training reduces chronic oxidative stress.
What is hormesis
Hormesis is a biological principle whereby:
A small stress activates protective adaptations that make the organism more resistant.
In exercise:
ROS transients → signal → adaptation → increased antioxidant capacity
Nrf2: the master regulator
Nrf2 (Nuclear factor erythroid 2–related factor 2) is a transcription factor that:
senses oxidative stress signals
migrates to the nucleus
activates antioxidant gene expression
Genes regulated by Nrf2 include:
glutathione synthase
glutathione peroxidase
superoxide dismutase
catalase
phase II detoxification enzymes
BOX — Key concept
Nrf2 does not neutralize ROS directly.
It increases the cell’s ability to defend itself.
What happens with regular training
Over time:
glutathione content increases
antioxidant enzymes increase
mitochondrial efficiency improves
basal ROS production decreases
Result: greater redox resilience.
Types of exercise and redox response
Aerobic
brisk walking
moderate running
cycling
Promotes:
mitochondrial biogenesis
Nrf2 activation
Strength
weights
bodyweight training
Promotes:
increased muscle mass
improved glucose metabolism
lower resting ROS production
HIIT
strong adaptive stimulus
useful if properly dosed
When exercise becomes harmful
Excess volume or intensity without recovery:
persistently elevated ROS
reduced immune function
increased inflammation
BOX — Optimal zone
Too little exercise → oxidative stress
Too much exercise → oxidative stress
Moderate dose → protective adaptation
Antioxidants and exercise: caution
High-dose vitamin C and E supplementation:
may blunt Nrf2 activation
may reduce some metabolic benefits of training
Integration with lifestyle
Exercise protection is maximal when combined with:
adequate sleep
balanced nutrition
stress management
Exercise as “medicine”
Physical activity:
reduces cardiovascular risk
improves insulin sensitivity
protects the brain
slows biological aging
Largely through improved redox balance.
BOX — Final key concept
Exercise does not reduce oxidative stress by eliminating ROS,
but by making the organism better able to handle them.
Conclusion
Physical exercise is one of the most powerful physiological tools for controlling oxidative stress.
Through transient ROS increases, it activates Nrf2 and triggers adaptations that strengthen endogenous antioxidant defenses, improving long-term cellular health.
✅ RELATED ARTICLE 4
Low-Grade Chronic Inflammation and Oxidative Stress
Highlight
Low-grade chronic inflammation is a persistent state of mild immune activation, often asymptomatic, that contributes to the development of many chronic diseases.
It is tightly intertwined with oxidative stress through a mutually amplifying circuit.
What is low-grade chronic inflammation
Unlike acute inflammation (rapid and resolving), it is:
persistent
systemic
low intensity
It does not cause obvious clinical signs but progressively alters tissue physiology.
Difference between acute and chronic inflammation
Acute inflammation
protective response
short duration
promotes healing
Low-grade chronic inflammation
continuous activation
lack of resolution
promotes tissue damage
BOX — Key concept
The problem is not inflammation itself, but its persistence.
Link with oxidative stress
Oxidative stress and inflammation form a bidirectional loop:
ROS activate inflammatory pathways
inflammatory cells produce ROS
BOX — Simplified circuit
ROS → cellular damage → inflammation → ROS production → further damage
Molecular mechanism
ROS activate transcription factors such as:
NF-κB
AP-1
These induce production of:
IL-6
TNF-α
other pro-inflammatory cytokines
Cytokines in turn increase:
oxidase activity
mitochondrial ROS production
Oxidative damage as primary event
Molecular damage caused by ROS can occur:
in absence of immune cells
directly to DNA, lipids, proteins
Inflammation represents a secondary response to damage.
BOX — Crucial point
Oxidative stress can initiate damage.
Inflammation maintains it.
Chronic inflammation and metabolism
Low-grade inflammation:
reduces insulin sensitivity
promotes dysfunctional lipolysis
increases ROS production
Explaining links with:
type 2 diabetes
metabolic syndrome
visceral obesity
Chronic inflammation and target organs
Involved in:
atherosclerosis
fatty liver
neurodegeneration
sarcopenia
Main factors promoting chronic inflammation
caloric excess
ultra-processed diet
sedentary lifestyle
sleep deprivation
psychological stress
gut dysbiosis
How to reduce chronic inflammation
Diet
high nutrient density
fiber
unsaturated fats
Physical activity
regular
Sleep
7–9 hours
Stress management
relaxation practices
BOX — Key concept
Reducing chronic inflammation also reduces oxidative stress.
Integration with other pillars
Inflammation is modulated by:
mitochondrial function
circadian rhythm
physical exercise
No single intervention is sufficient.
Conclusion
Low-grade chronic inflammation and oxidative stress form an integrated system of biological damage amplification.
Interrupting this circuit requires a systemic approach acting on metabolism, lifestyle, and neuroendocrine regulation.
✅ RELATED ARTICLE 5
Biomarkers of Oxidative Stress: What to Measure and How to Interpret
Highlight
Oxidative stress cannot be evaluated with a single test.
A clinically meaningful assessment requires integration of biomarkers of oxidative damage, inflammation, antioxidant capacity, and metabolic context.
Why there is no “perfect marker”
Oxidative stress is a dynamic process involving:
ROS production
molecular damage
antioxidant response
repair
Each biomarker observes only one part.
BOX — Key concept
A panel is more informative than a single value.
1) Direct biomarkers of oxidative damage
F2-isoprostanes
Derived from non-enzymatic lipid peroxidation
Considered gold standard for lipid oxidative damage
Sample: plasma or urine
Interpretation:
High → high lipid oxidative stress
Malondialdehyde (MDA)
Lipid peroxidation product
More variable than isoprostanes
Interpretation:
Useful as orientative indicator
8-OHdG (8-hydroxy-2’-deoxyguanosine)
Marker of oxidative DNA damage
Urine or blood
Interpretation:
High → increased DNA oxidation
2) Antioxidant capacity biomarkers
Reduced glutathione (GSH) and GSH/GSSG ratio
Central redox parameter
Interpretation:
High ratio → good balance
Low ratio → oxidative stress
Total antioxidant capacity (TAC)
Global estimate of ROS-neutralizing ability
Low specificity
Interpretation:
Useful as complement
3) Inflammation-related biomarkers
hs-CRP
Integrated marker of systemic inflammation
Indicative values:
<1 mg/L → low CV risk
1–3 mg/L → intermediate risk
3 mg/L → high risk
IL-6, TNF-α
Pro-inflammatory cytokines
Mainly specialist use
4) Indirect metabolic biomarkers
Glucose, insulin, HOMA-IR
Triglycerides, oxLDL
Ferritin
BOX — Key concept
Metabolic alterations are often the main source of chronic oxidative stress.
5) Advanced mitochondrial biomarkers
Resting lactate
Lactate/pyruvate ratio
CoQ10
Useful in specialist settings.
6) Minimal practical panel
hs-CRP
F2-isoprostanes or MDA
8-OHdG
GSH/GSSG
Glucose + insulin
7) Integrated interpretation example
hs-CRP ↑
MDA ↑
GSH/GSSG ↓
Indicates:
active oxidative stress
associated inflammation
reduced defenses
8) Temporal changes after intervention
Improve first:
GSH/GSSG
hs-CRP
Later:
MDA / F2-isoprostanes
Slowest:
8-OHdG
BOX — Typical sequence
Protection rises → damage falls → DNA improves
9) Common errors
Relying on one marker
Using TAC alone
Interpreting without clinical context
Conclusion
Assessment of oxidative stress requires a multiparametric approach.
Integrating damage, antioxidant capacity, inflammation, and metabolism allows a biologically coherent reading of redox status.
✅ RELATED ARTICLE 6
Antioxidant Supplements: When They Are Truly Needed
Highlight
Antioxidant supplements are not a universal solution to oxidative stress.
In many cases, indiscriminate use is useless or potentially counterproductive.
The most effective strategy remains strengthening endogenous antioxidant defenses.
Why “more antioxidants = less ROS” is wrong
ROS:
are not only toxic byproducts
have essential physiological functions
Indiscriminately eliminating ROS can:
interfere with signaling
reduce beneficial adaptations
BOX — Key concept
The goal is not to suppress ROS, but to restore redox balance.
What dietary antioxidants really do
Dietary antioxidants:
partially buffer ROS
mainly activate signaling pathways (e.g., Nrf2)
Many polyphenols act more as adaptive signals than direct scavengers.
Evidence on high-dose supplements
Chronic high-dose vitamin C and E:
may reduce exercise metabolic benefits
may blunt Nrf2 activation
When supplementation may be useful
Documented deficiencies
vitamin C
vitamin E
selenium
zinc
Increased demand
high stress
infections
toxin exposure
recovery phases
Specific clinical conditions
malabsorption
selected chronic diseases
Types of integrative approach
Direct antioxidants
vitamin C
vitamin E
Glutathione precursors
N-acetylcysteine
glycine
Mitochondrial modulators
CoQ10
alpha-lipoic acid
BOX — Preferred strategy
Better to provide substrates and signals to produce endogenous antioxidants than large doses of external scavengers.
Risks of abuse
reduced training adaptations
possible increased mortality in some populations
false sense of security delaying lifestyle change
Correct intervention sequence
Sleep
Nutrition
Physical activity
Stress management
Only then: targeted supplementation
Supplementation and personalization
Good supplementation:
is temporary
is biomarker-based
is re-evaluated
Conclusion
Antioxidant supplements do not replace a healthy lifestyle.
They may play a targeted role in selected contexts, but the most effective protection against oxidative stress comes from strengthening the body’s intrinsic capacity.