“Una volta ragionevolmente raggiunta la diagnosi di NCGS, la gestione e il follow-up dei pazienti sono completamente oscuri. Un approccio logico consiste nell’intraprendere un regime dietetico senza glutine per un periodo limitato (ad esempio, sei mesi), seguito dalla graduale reintroduzione del glutine. Durante la dieta priva di glutine, dovrebbe essere evitata l’ingestione del peptide prolaminico (gliadina), derivato da grano, segale, orzo, avena, bulgur e ibridi di questi cereali. Riso, mais e patate sono stati i tipici sostituti, ma oggigiorno si possono usare altri cereali e pseudocereali diversi come amaranto, grano saraceno, manioca, fonio, teff, miglio, quinoa e sorgo. Dopo un po ‘di tempo con una dieta priva di glutine, la reintroduzione del glutine può iniziare con cereali a basso contenuto di glutine (ad es. Avena). Inoltre, è possibile utilizzare il farro einkorn (Triticum monococcum), che non presenta tossicità diretta in vitro o ex vivo e basso contenuto di glutine (7%)[41]”. (Non-celiac gluten sensitivity: Time for sifting the grain. Luca Elli, Leda Roncoroni, and Maria Teresa Bardella. Copyright ©The Author(s) 2015. Published by Baishideng Publishing Group Inc. All rights reserved).
Prevenzione della celiachia: dieta con grano con ridotta quantità di epitopi stimolatori delle principali T-cellule.
La ricerca scientifica ha più volte sostenuto ed incoraggiato l’utilizzazione di grani a bassa tossicità nella prevenzione della celiachia; Nella ricerca che ora presentiamo sono stati studiati alcuni grani evidenziando il loro profilo riguardo sia la presenza dei peptidi resistenti alla digestione gastro-intestinale sia, tra questi, quelli contenenti la frazione “tossica” (Summary of the GD-resistant peptides identified at the end of the duodenal phase and counting of the peptides encrypting full length epitopes relevant for celiac disease (CD) and wheat allergy (table 3) “….omissis Even if none of them can be considered safe for CD patients, grain with reduced amount of major T-cell stimulatory epitopes may help in the prevention of CD, since previous studies demonstrated that the amount and duration to gluten exposure are strictly linked to the initiation of this pathology.” (A Comprehensive Peptidomic Approach to Characterize the Protein Profile of Selected Durum Wheat Genotypes: Implication for Coeliac Disease and Wheat Allergy. Rosa Pilolli , Agata Gadaleta, Luigia Di Stasio , Antonella Lamonaca, Elisabetta De Angelis , Domenica Nigro , Maria De Angelis , Gianfranco Mamone and Linda Monac. Published: 1 October 2019).
Abstract
The wheat varietal selection undertaken by breeders in recent decades has been tailored mainly to improve technological and productivity-related traits; however, the latter has resulted in a considerable impoverishment of the genetic diversity of wheat-based products available on the market. This pitfall has encouraged researchers to revalue the natural diversity of cultivated and non-cultivated wheat genotypes in light of their different toxic/immunogenic potential for celiac disease and wheat-allergic patients. In the present investigation, an advanced proteomic approach was designed for the global characterization of the protein profile of selected tetraploid wheat genotypes (Triticum turgidum). The approach combined proteins/peptides sequence information retrieved by specific enzymatic digestions (single and dual proteolytic enzymes) with protein digestibility information disclosed by means of in-vitro simulated human gastroduodenal digestion experiments. In both cases, the peptide pools were characterized by discovery analysis with liquid chromatography high-resolution tandem mass spectrometry, and specific amino acid sequences were identified via commercial software. The peptide list was screened for in silico toxicity/immunogenicity risk assessment, with the aid of various open-source bioinformatics tools for epitopes matching. Given the global information provided by the designed proteomic approach, the in silico risk assessment not only tackled toxicity implication for celiac disease patients, but also scouted for immunogenic sequences relevant for wheat allergic patients, achieving a comprehensive characterization of the protein profile of the selected genotypes. These latter were assessed to encrypt a variable number of toxic/immunogenic epitopes for celiac disease and wheat allergy, and as such they could represent convenient bases for breeding practices and for the development of new detoxification strategies.
La genetica della celiachia
Prevenzione della celiachia
Caratterizzazione di varietà autoctone di grano duro siciliane, utilizzando marcatori di polimorfismi a singolo nucleotide (forniscono un’impronta genetica molto utile per evitare frodi commerciali).
Riassunto: Nel corso del XX secolo, l’uso diffuso delle moderne cultivar di grano ha ridotto drasticamente la coltivazione di antiche varietà autoctone, che oggi sono confinate in aree di coltivazione di nicchia. Diverse varietà autoctone di grano duro adatte agli ambienti estremi della regione mediterranea, sono ancora coltivate in Sicilia, in Italia. Una conoscenza dettagliata della diversità genetica di questo germoplasma potrebbe gettare le basi per la loro gestione efficiente nei programmi di riproduzione, per una vasta gamma di caratteristiche. Lo scopo del presente studio era di caratterizzare una raccolta di varietà autoctone di grano duro siciliane, utilizzando marcatori di polimorfismi a singolo nucleotide (SNP), insieme a tratti agro-morfologici, fenologici e relativi alla qualità. Due grani moderni cv. Simeto, Claudio e uno autoctono esaploide, Cuccitta, furono usati come outgroup. L’analisi dei cluster e l’analisi delle coordinate principali (PCoA) ci hanno permesso di identificare quattro cluster principali attraverso il germoplasma analizzato, tra cui un cluster includeva solo varietà storiche e moderne. Allo stesso modo, l’analisi della struttura è stata in grado di distinguere le varietà antiche dalle altre, raggruppando le voci in sette cluster genetici criptici. Inoltre, un’analisi delle componenti principali (PCA) è stata in grado di separare i moderni tester dall’antico germoplasma. Questo approccio è stato utile per classificare e valutare il germoplasma di grano antico siciliano, supportando la loro salvaguardia e fornendo un’impronta genetica necessaria per evitare frodi commerciali per sostenere i profitti economici degli agricoltori che ricorrono alla coltivazione delle terre. (High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Maria Carola Fiore , Francesco Mercati, Alfio Spina, Sebastiano Blangiforti, Gianfranco Venora, Matteo Dell’Acqua, Antonio Lupini, Giovanni Preiti , Michele Monti, Mario Enrico Pè and Francesco Sunseri. 30 aprile 2019.)
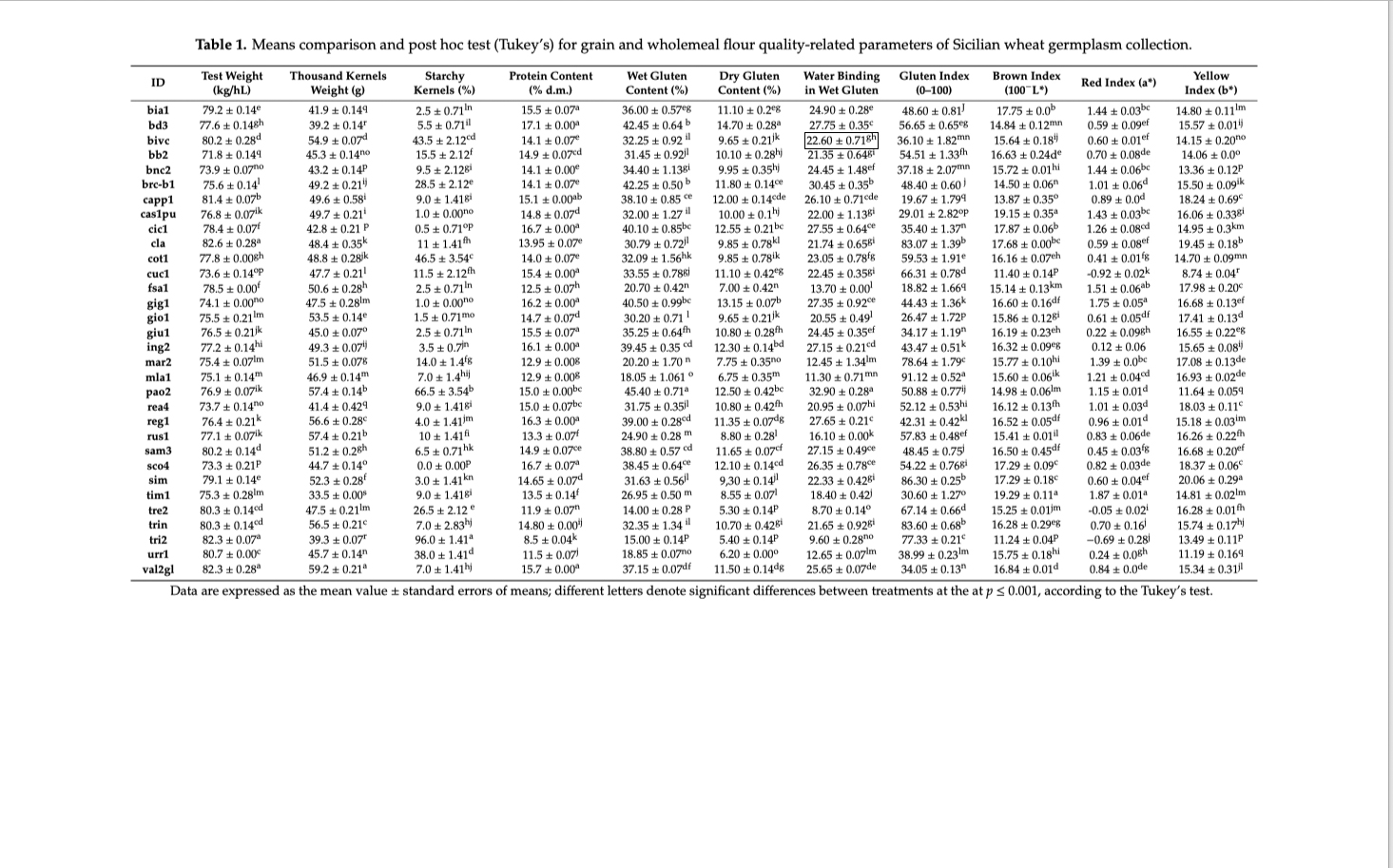
Tavola 1
Lista grani esaminati
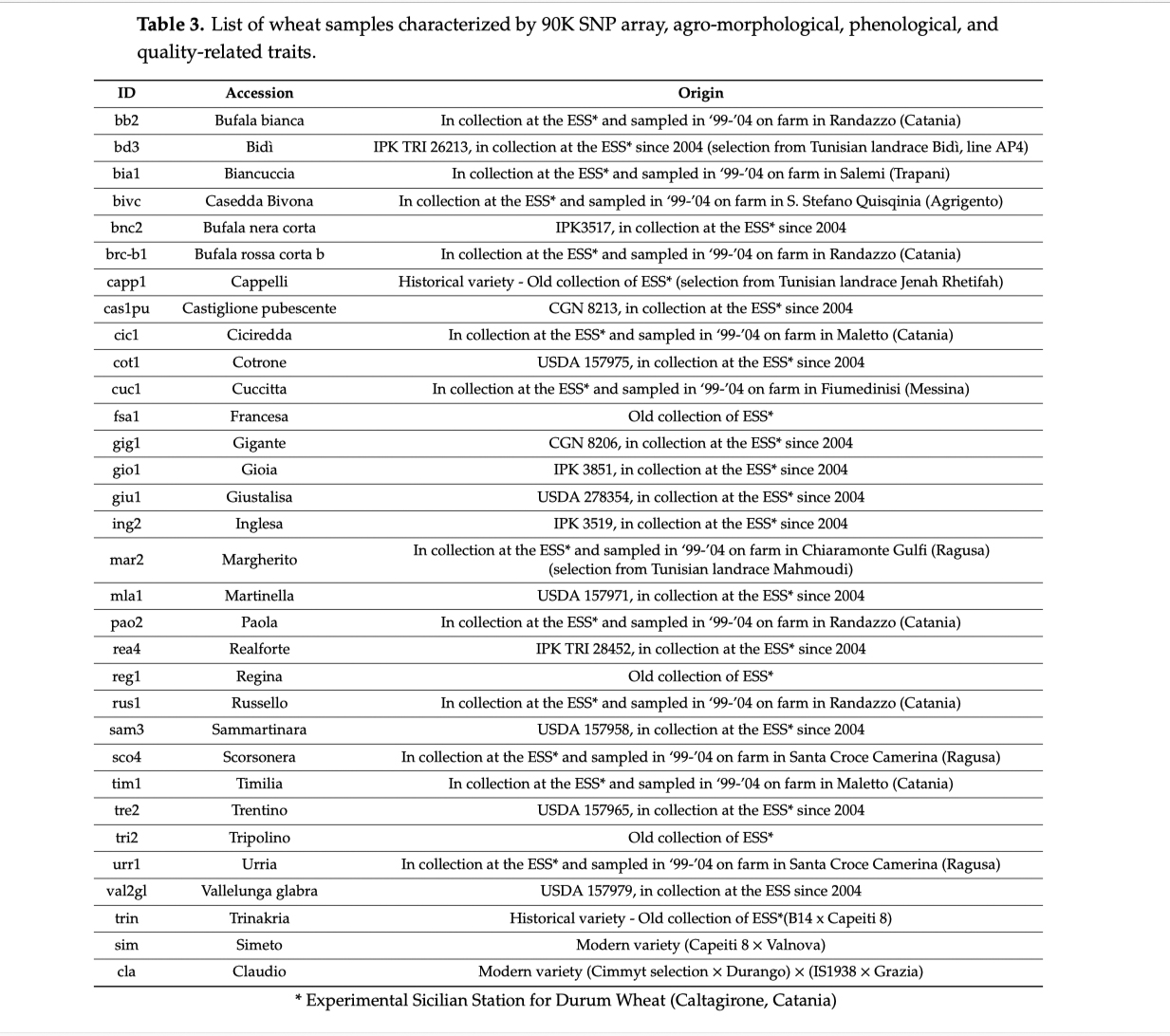
Tavola 2
Caratteristiche dei grani esaminati